�l���r�g��2022-01-26���ٷ���r�IՓ���g�[��1��
ժ Ҫ�� ժ Ҫ�����ýM�����x�����x���Kʡ���w�ж���ֲ���������˵ĺڸ�����ԭ����ͨ�^�ΑB�W�^�졢rDNAITS ���з������²��Ԝy�����_����ԭ���ķ�W��λ�����y���ضȡ�pH�����ա�̼Դ�͵�Դ����ԭ�����z���L�ͮa������Ӱ푡��Y���������������w�^�������˺ڸ������²���ԭ
����ժ Ҫ�����ýM�����x�����x���Kʡ���w�ж���ֲ���������˵ĺڸ�����ԭ����ͨ�^�ΑB�W�^�졢rDNA–ITS ���з������²��Ԝy�����_����ԭ���ķ�W��λ�����y���ضȡ�pH�����ա�̼Դ�͵�Դ����ԭ�����z���L�ͮa������Ӱ푡��Y���������������w�^�������˺ڸ������²���ԭ�������_̿�Ҿ�(Colletotrichum siamense)��ԓ��ԭ�����m���z���L�ͮa�����ĜضȞ� 30 �棬5 ��r���z���L�O�侏����ֹͣ�a�ߣ�40 ��r���z������ֹͣ�a��;���z���L�����m pH 5.0���a�ߵ����m pH 7.0;��ԭ�������ö�N̼Դ�͵�Դ����ѿ�Ǟ���z���L�ͮa�ߵ����̼Դ�������˞�a����ѵ�Դ����ĸ��������z���L��ѵ�Դ�������@�����z���L�ͮa������������;�ⰵ�����m�Ͼ��z���L���B�m�ڰ��m�Ϯa�ߡ�
�����P �I �~����������;�ڸ���;���_̿�Ҿ�;����W����
��������ֲ����������(Pachyphytum ‘Dr Cornelius’) �Ǿ����(Crassulaceae)���~��(Pachyphytum)�͔Mʯɏ����(Echeveria)�Č��g�s���N��ԭ�a��ī����[1]���ǽ��K���w�Nֲ����Ҫ����ֲ��Ʒ�N����������������˺ڸ����l���^����أ��ѳɞ����w����ֲ��a�^����Ҫ����������Ӱ��������˵�Ʒ�|�ͷNֲҎģ����������ֲ��ĸ����o���~�����Ȳ�λ������Ⱦ�����~Ƭ�ܺ�������ɫ��׃ܛ��ˮ������ή��Ȼ�������~ƬѸ�ٰl�ڸ�����������ֲ����������;��ˣ����_�������˺ڸ���ԭ���N�������W���ԣ����������˺ڸ����ķ��ؾ�����Ҫ���x��Ŀǰ�����P����ֲ��ڸ�����ԭ�YՓ��һ����ؐؐ[2]����ɽ�|���ݵ^����ƶ���ֲ��ѩɏ�Ϸ��x���ڸ����IJ�ԭ���� Fusarium inflexum ;���Ƶ�[3]�о��J�飬����ֲ��ʺ�ڸ�����ԭ���� Fusarium oxysporum;Ҧ�\�۵�[4]�J�鸣�����ݾ���ƶ���ֲ����侰��ڸ�����ԭ����ɽ�ⶹ������(Corynespora cassiicola)����Ҋ����ͬ����ֲ��ĺڸ������ɲ�ͬ��ԭ����Ⱦ��ɵġ��P�ߏĽ��Kʡ���w�й�܇悶���ֲ��Nֲ���زɼ��������˵ĺڸ������Ͳ��꣬���x��������ԭ���������ΑB�W�^�졢rDNA–ITS �������з������²��Ԝy����Y�ϵķ����M�в�ԭ�b����������ԭ���M������W�����о������ڞ����ֲ���������˺ڸ����ķ����ṩ�ƌW������
����1 �����c����
����1.1 ����
������ 2019 �� 8 �²ɼ����w�й�܇悶���ֲ��Nֲ���ص��������˺ڸ������ͰY��IJ����˱������������w�WԺ�@���Iֲ�ﲡ�팍��ҡ�
����1.2 ����
����1.2.1 �������˺ڸ�����ԭ���ķ��x���b��
���������īI[5]�ķ����������҃ȽM�����x���@���������˺ڸ�����ԭ���꣬���� 26 �� ������ PDA б�����B�������á�
��������ԭ���ӷN�� PDA ƽ���ϣ����B 5 d���^������ΑB���ɫ�����z���L��r;���B 10 d ������W�@�R���^��������Ӻ;��z���������ΑB�����������������M DNA ��ȡԇ����(�Ͼ��������Ƽ�����˾�aƷ����̖ D2300–50T)��ȡ��ԭ����Ļ���M DNA���� rDNA–ITS ����ͨ������ ITS1/ITS4(ITS1��5'–TCCGTAGGTGAACCTGCGG– 3';ITS4��5'–TCCTCCGCTTATTGATATGC–3')PCR �U���������l����95 ���A׃�� 5 min;95 ��׃�� 15 s��50 ���˻� 20 s��72 ������ 40 s��40 ��ѭ�h����� 72 ������ 5 min���� Axygen DNA ���z����ԇ����(Ŀ�̖ AP–GX–50)�f������������ PCR �U���a������Ͼ���˹������Ƽ�����˾�y�����@�õĻ������У��� NCBI �Wվ�M�� BLAST ͬԴ�ԱȌ���ͨ�^�Ȍ��xȡ���ƶȸߵ�20 �l�������У����� MEGA 6.0 ܛ���M��ϵ�y�l����Ę�����
����1.2.2 ��ԭ���²��ԵĜy��
�������x�����ڼs 1 �ꡢ��� 12~15 cm �Ľ���������������ֲ�������²��Ԝy�����xȡ��������ֲ���в����~���ßo��ˮ���͛_ϴ���� 75%���Ҵ����� 3 s���� 0.1%�� HgCl2��Һ���� 3 min�����ßo��ˮ�_ϴ 3~4 �Σ�����Ü������ˮ�����ɱ����ˮ�֡��ßo����^�p�p�̂�����̎���^�����������~Ƭ�ϱ�Ƥ���ڂ���̎���� 3 mm×3 mm ���y����ľ��z�K���ԟo�� PDA ��֬�K�ӷN�����ա����ӷN�����������~Ƭ���� 26 ���ֲ������ȣ�12 h ���ռ� 12 h �ڰ�̎�����^��l����r�����l���ڸ������ͰY������������~Ƭ���M���ٴη��x�����M���ΑB�W�^��ͷ����b����
����1.2.3 ��ԭ������W���ԵĜy��
�����x�� PDA ƽ�壬pH �� 7.0���O�� 5��10��15�� 20��25��30��35��40 �� �� 8 ���ض��ݶȣ�3 ���؏͡��Ü�������ȡֱ���� 5 mm �ľ��P������ƽ�������룬����ȫ�ڰ��l�������B 5 d��ÿ������y�� 3 ������ֱ����ȡƽ��ֵ��Ӌ����z���L���ʣ����B 10 d ��ÿ����� 10 mL �o��ˮϴÓ���ӣ���Ѫ��Ӌ���巨[6]�y���a������
������ 1 mol/L �� HCl �� 1 mol/L �� NaOH �� PDA ���B���քe�{���� pH �� 3.0��4.0��5.0��6.0��7.0�� 8.0��9.0��10.0��11.0���� 9 ��̎����3 ���؏ͣ��� 26 ���y�����z���L���ʺͮa������
�����x�� PDA ���B����pH �� 7.0���O�� 24 h �B�m�ڰ���24 h �B�m���պⰵ����(12 h ���ռ� 12 h�ڰ�)3 ��̎����3 ���؏ͣ��� 26 ���y�����z���L���ʺͮa������
�������պ�������[7]�ķ�������������������B�� (MgSO4·7H2O 0. 5 g��KH2PO4 0. 5 g����֬ 20 g��̼Դ 5 g����Դ 1 g�����sˮ 1000 mL)���քe��ľ�Ǵ������ǡ������Ե��ۡ�D–���ǡ������Ǻ���ѿ�Ǟ�̼Դ���Բ���̼Դ���B���錦��;�քe�����ء���ĸ���ࡢ�����@�������ˡ��ʰ��������鵪Դ���Բ��ӵ�Դ���B���錦�գ��� 26 ���y�����z���L���ʺͮa������
����2 �Y���c����
����2.1 �������˺ڸ�����ԭ�����b���Y��
�����������˺ڸ�����ԭ���麦ֲ��������o�����~Ƭ���o�����~Ƭ�l���^�ء��~Ƭ���Ļ����_ʼ�l�������~�˾G��ܛ������u��ˮ���l����λ�_ʼ���F�ڰߣ�������u�U����ȫ�~���l�������~Ƭ�ϳ��F�ں�ɫС�c(����)������Ó��(�D 1)��
�������x�������������B�Ԡ���D 2 ��ʾ����������(�D 2–1)�a���\�Ұ�ɫ�������z�����z�l�_���^������;���䱳��(�D 2–2)��\�Ұ�ɫ�������������Ԏ����Sɫ;���z������(�D 2–3)���غ�ɫ�����A�λ�E�A�ͣ�߅��������ƽ����Ҏ�t;��������(�D 2–4)���A���һ���g�A����һ���g�A���Լ⼚��������ԭ�����ΑB�W�������Y�ϗ�����[8]�����[9]�����ܵ�[10]���쵤����[11]�� SOARES ��[12]�������������ж�ԓ������܌����z��̿�ҏͺϷN (Colletotrichum gloeosporioides species complex)�����_̿�ҏͺϷN(Colletotrichum siamense species complex)�� 1 ��������;2 ���䱳��;3 ���z������;4 �������ӡ��D 2 �������˺ڸ�����ԭ�����ΑB���� Fig.2 The morphological characteristic of black rot pathogen in Pachyphytum ‘Dr Cornelius’ �Է��x�õ��IJ�ԭ���� DNA ��ģ�壬����ͨ������ IST1 �� IST4 �M�� PCR �U�����õ��� ITS ����Ӿ����С�� 500 bp ���ϡ��y��Y��������ԓ PCR �a��U����Ŀ�Ļ���Ƭ������ȫ�L�� 543 bp���� NCBI/BLAST �ό�ԓ����(J257–01)�Ļ��������M��ͬԴ�ԱȌ����l�F���c���_̿�Ҿ� C. siamense ģʽ��������(Gene accession No. KM268865.1�� KC702973.1�� MH939973.1��KX786431.1��MH939974.1�� MH939977.1��MT229430.1)���ƶ��_ 100%;�c fungal sp. ģʽ�������� (Gene accession No. MG490812.1��MG490789.1�� MG490768.1��MG490759.1) ���ƶȞ� 99%;�c C. tropicale ģʽ��������(Gene accession No. MK330040.1)���ƶȞ� 99%���� NCBI/ GenBank ���xȡ��Դ���� rDNA–ITS �������У��� Colletotrichum parsonsiae ����(Gene accession No. MH865006.1)������Ⱥ���И���ϵ�y�l���䣬�Y��(�D 3)�������xȡ�Ľ�Դ DNA ���з��� 4 ����ͬ���M����֧��C. siamense �c C. tropicale �H���Pϵ���^�������M���ό���ͬһ��֧�������������˺ڸ����ľ��� PCR �U���a���c C. siamense ����ͬһ��֧�����Դ_�������������˺ڸ����ľ�������_̿�Ҿ�(Colletotrichum siamense)��
����2.2 �������˺ڸ�����ԭ�����²���
���������������ľ��z�ӷN�ڽ��������������~Ƭ�������^�y�l����r���D 4–1 �錦�գ���Ҋ�~Ƭ�Ա��־Gɫ���~Ƭ���l�����H�ڽӷN����̎���F�ں�ɫ���������c���S���r�g�����ӣ��������c�]�����D 4–2 �͈D 4–3 ��Ӿ��~Ƭ���Ӿ� 2 d ���~Ƭ���S�����Fˮ���F�Ӿ���̎�_ʼ���F�ڰߣ�����ֱ�� 3.0~3.5 mm;�Ӿ� 5 d ���~Ƭ׃������ȫ��ˮ���ڰ߸��w�����~Ƭ���~Ƭ�ϳ��F�ں�ɫС�c(����)�����ӷN�l�������������~Ƭ�ٴ��M�з��x�b�����@�õľ����cԭ�ӷN����һ�£����ж��������˺ڸ������²��������_̿�Ҿ� (C. siamense)��
����2.3 �������˺ڸ�����������W����
����2.3.1 �ضȡ�pH ���շ�ʽ�����z���L�ͮa�ߔ�����Ӱ�
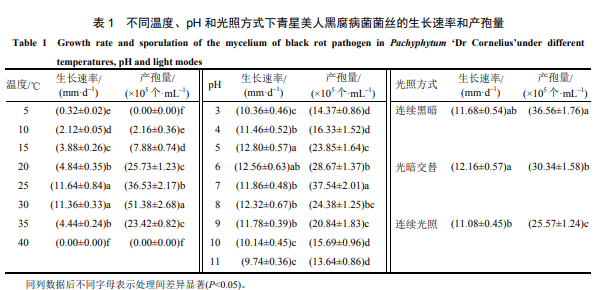
�����ɱ� 1 ��֪����ͬ�ضȌ��������˺ڸ������ľ��z���L�ͮa������Ӱ푣�5~35 ��r�����z�������L��25 ��� 30 ��r���z���L�����^�քe�� 10 �� �r�ľ��z���L��������� 449.06%�� 435.85%���@�����������ض��µľ��z���L����;10~35 ��r���������ܮa�ߣ����m�a�ߜضȞ� 30 �棬��a������ 10 ��r�Įa���������� 2278.70%���@�����������ض��µĮa������5 ��r�����z���L�O�侏������ 10 �� �r�ľ��z���L���ʜp���� 84.91%��ֹͣ�a��;40 �� �r�����z������ֹͣ�a�ߡ�
�������z�� pH3~11 �����B���Ͼ����^�����L��pH ֵ�� 11 �r�����z���L������С��pH ֵ�� 5 �r���z���L������� pH �� 11 �r�ľ��z���L��������� 31.42%���c pH ֵ�� 6 �r�ľ��z���L���ʟo�@��������@���������� pH �µľ��z���L���ʡ�pH ֵ�� 11 �r�a������С��pH ֵ�� 7 �r�a������ߣ��� pH11 �r�Įa���������� 175.22%���@���������� pH �µĮa������
�����B�m���շ�ʽ�¾��z�����L���ʺͮa������ͣ��ⰵ���淽ʽ�¾��z���L��������B�m���շ�ʽ�µľ��z���L���������� 9.75%�����c�B�m�ڰ��µľ��z���L���ʟo�@���;3 �N���l���µĮa���������@������B�m�ڰ����m�Ϯa�ߣ��a�������B�m���շ�ʽ�µĮa��������� 42.98%��
����2.3.2 ̼Դ�͵�Դ�����z���L�ͮa������Ӱ�
�����ɱ� 2 ��֪����̼ͬԴ���B���ϵľ��z���L���ʾ��@�����ڌ��գ�����ѿ�Ǻ����Ǟ�̼Դ�ľ��z���L�����^�քe�Ȍ��������� 40.70%�� 38.67%���@������ľ�Ǵ��������ǣ��c�����Ե��ۺ� D–����֮�g�o�@���;��̼ͬԴ���B���ϵĮa�������@�����ڌ��գ���ѿ�Ǟ����m�a��̼Դ���a�����Ȍ��������� 71.36%���@����������̼Դ��
�����������@�鵪Դ�ľ��z���L���ʺͮa�����@��С�ڌ��գ��քe�Ȍ��՜p���� 17.29%�� 19.94%;��ĸ��������z���L���m��Դ������z���L���ʱȌ��������� 32.82%���c������֮�g�o�@��������@������������Դ;�����˞����m�a�ߵ�Դ����a�����Ȍ��������� 60.31%���@������������Դ�ġ�
����3 �YՓ��ӑՓ
�������о��Y�����������Kʡ���w�й�܇悶���ֲ��Nֲ�����������˺ڸ����²��������_̿�Ҿ�(C. siamense)��ԓ������Ҫ�l���ڞ� 5—9 �£���ԭ������A�h�����m���Զ��^�����B�m���տ���Ч���Ʋ����l���������@�����z���L�ͮa�����������á�
�������_̿�Ҿ����z��̿�Ҿ�(C. gloeosporioides) �ͺϷN֮һ�������� 2009 ����̩�������ϱ��l�F�������l����̿�Ҳ�[13]���S����������_̿�Ҿ���Ⱦֲ�����l���������P������������[8]�J�飬���_̿�Ҿ���������ˮ����ժ���̿�Ҳ�;��������[14] �J�飬���_̿�Ҿ��������Z�Ʋ�̿�Ҳ�;��ӵ�[15] �J�飬���_̿�Ҿ��������Ͳ�����̿�Ҳ�;�n���µ�[16]��������_̿�Ҿ���������ɽˎ̿�Ҳ������Ї���W�߈�����_̿�Ҿ��������ͩ��ɽˎ��������ֲ���̿�Ҳ�[17–19]�����@Щ�����֪���_̿�Ҿ���Ⱦֲ������IJ�����̿�Ҳ������P�߰l�F���_̿�Ҿ���Ⱦ����ֲ���������˿����l�ڸ���������Ƽ��[20]�J�飬̿�Ҿ��ٲ�ԭ�����²��^�����γɸ�������ͬ�r�����ϳ�һ�N�������x���|——��ɫ�ء�
������ɫ�س���Ӱ푲������²����⣬߀��ʹֲ���~Ƭ�ڻ�[21–22]���_����[23]�l�F�����_̿�Ҿ��܉�����ƣ�}��‘����̖’���ٸ�����������Ƽ��[20]�J�飬̿�Ҿ��ٲ�ԭ������ڰ���w�I�B����������������ֲ��ǰ�ڣ�������������ֲ��I���w�I�B����;�����ֺ��ڣ��S����Ⱦ���z�����ӣ�����ֲ��I���w�I�B�������ɴ˿�֪�����_̿�Ҿ�����Ⱦֲ��������ֲ��l�ڡ��������������������_̿�Ҿ�����Ⱦ�������˳����~Ƭ�S�����S��������ֲ���w�ȴ�����ֳ�����~Ƭ�_ʼ���F�l�ڡ������������ȰY�Ҧ�\�۵�[4]�о�������������ֲ���~Ƭ�ְ߲���ɽ�ⶹ�����߾�Ҳ�����l����ֲ��ĺڸ�����ԓ�Y���c���о��Y�������ڲ�ԭ����Ⱦ����ֲ������ǵ����Y����������@�����c����ֲ���~Ƭ�ʺ����|��֭�Һ�ˮ��������c���P�����_̿�Ҿ���Ⱦ�����������l�ڸ������²��C��߀��Ҫ�Mһ���о���
�������о��Y���@ʾ��10~35 ��r���������˺ڸ�����ԭ�����_̿�Ҿ����z���L�ͮa�߾�������5 �� �r���z���L�O�侏����ֹͣ�a�ߣ�40 ��r���z������ֹͣ�a�ߡ����m���z���L�ͮa�ߵĜضȞ� 30 �棬�@�c������[24]���о��Y��һ�£��f���������˺ڸ������m�İl����؞� 25~30 �棬���Kʡ���w 5—9 �µĚ��ǡ���c�������˺ڸ����l���m�˚����ϡ��������˺ڸ�����ԭ�����_̿�Ҿ��� pH ���m�������^�V��pH ֵ 3~11 �r�����z���L���ã�pH ֵ 5 �r�����z���L�������;pH ֵ 5~9 �r���a�����^��pH ֵ 7 �r���a�������ԓ�Y���c���[9] �ĺ���ʡ�Ͳ輰������ֲ���Ϸ��x�������_̿�Ҿ��� pH ֵ 4~11 ���z���L���õĽY�����ƣ���Ҋ�������˺ڸ���������A�h�����m�����^�����m���������˺ڸ�����ԭ�����_̿�Ҿ����z���L�ͮa�ߵ����̼Դ����ѿ�ǣ���ѵ�Դ�鵰���ˣ������@�����z���L�ͮa�����������ã��ⰵ�����m�Ͼ��z���L���B�m�ڰ����m�ϲ�ԭ���a�ߣ���������[24] �о��J�飬�m�Z���̿�Ҳ���ԭ�����_̿�Ҿ����m���z���L�ͮa�ߵ�Դ�鵰���ˣ����m���z���L̼Դ�� α–���ǣ���Ѯa��̼Դ�������ǣ��B�m�ڰ��m�Ͼ��z���L���ⰵ�����m�Ϯa�ߣ��@��������鲻ͬ����ֲ�ꡢ��ͬ�����Դ��ͬ�N��������B�m���Բ�ͬ[25]�ľ��ʡ�——Փ�����ߣ����������������n���£����f�������A
���������īI��
����[1] �����������ϣ��w������.�������������~ƬǤ�己ֳ�wϵ��̽[J].�F���@ˇ��2019(21)��24–26. ZHANG J Y��ZHU Y��ZHAO W��et al.Preliminary study on the leaf cutting propagation system of succulent plant Pachyphytum ‘Dr Cornelius’[J].Xiandai Horticulture�� 2019(21)��24–26.
����[2] ��ؐؐ.������H�������ĺY�x������M�y�����B������[D].̩����ɽ�|�r�I��W��2018. WANG B B . Screening �� genome sequencing and optimization of medium and of PGPR from the rhizosophere of Crassulaceae plants[D].Tai’an��China�� Shandong Agricultural University��2018.
����[3] ���ƣ���ˬ��������.����ֲ��ʺ�ڸ�����ԭ���ķ��x�b��[C]//������.�Ї�ֲ������W�� 2019 ��W�g���Փ�ļ�.�������Ї��r�I�ƌW���g�����磬 2019��199. LIU H��YANG S��TIAN P Y��et al.Isolation and identification of the pathogen of succulent rainbow black rot[C]//PENG Y L.Proceedings of the 2019 Annual Meeting of the Chinese Society of Plant Pathology. Beijing��China Agricultural Science and Technology Press��2019��199.
����[4] Ҧ�\�ۣ��S�i��ꐝh�Σ���.����ֲ����侰��ڸ�����ԭ��[J].����W��2020��39(2)��452–456. YAO J A��HUANG P��CHEN H X��et al.Black rot pathogen of succulent plant Sedum morganianum[J]. Mycosystema��2020��39(2)��452–456.
����[5] �����_.ֲ���о�����[M].3 ��.�������Ї��r�I�����磬1998. FANG Z D.Plant Disease Research Method[M].The third edition.Beijing��China Agricultural Press��1998.
����[6] �����壬��������������.���~��̿�Ҳ���ԭ�����b���c����W����[J].�㽭�r�I�W��2020��32(11)�� 2009–2019. LI X Q��YAN J L��RUAN S L.Identification and biological characteristics of anthracnose pathogen on Tetrastigma hemsleyanum[J].Acta Agriculturae Zhejiangensis��2020��32(11)��2009–2019.
SCISSCIAHCI