�l���r�g��2022-04-12���ٷ���r�IՓ���g�[��1��
ժ Ҫ�� ժ Ҫ ��Ŀǰ GenBank �����칲��� 167 �Nֱ��Ŀ���xȫ�����w����M���У��漰�ȁ�Ŀ 9 ������ 22 ���� 99 ����N��Ŀ 7 ������ 12 ���� 68 ����N���ڴ˻��A�ϣ�ԓ�ķ�����ֱ��Ŀ���x�����w����M�Ļ��������������˾����wȫ����M��ֱ��Ŀ���xϵ�y�l���о��ϵ�
����ժ��Ҫ ��Ŀǰ GenBank �����칲��� 167 �Nֱ��Ŀ���xȫ�����w����M���У��漰�ȁ�Ŀ 9 ������ 22 ���� 99 ����N��Ŀ 7 ������ 12 ���� 68 ����N���ڴ˻��A�ϣ�ԓ�ķ�����ֱ��Ŀ���x�����w����M�Ļ��������������˾����wȫ����M��ֱ��Ŀ���xϵ�y�l���о��ϵđ��� ;ͬ�r���ھ����wȫ����M�����ؽ���ֱ��Ŀ���x��ϵ�y�l���Pϵ����Ҫ�Y������ ��(1) ֱ��Ŀ���x���� 8 �N�����w����M������ͣ����� trnK-trnD ���ŬF��H�l���ڻȿ����У�trnN-trnS-trnE ���ŬF��H�l��������У�trnM-trnI-(-trnQ) ���ŬF��H�l���ڔM�~�x������ ;(2) ֱ��Ŀ���xȫ�����w����M�ĉA���M�ɾ������@�� AT ƫ���� ;(3) ��ͬ�ĵ����|���a������ֱ��Ŀ���x�е��M�����ʲ�ͬ ;(4) ֧��ֱ��Ŀ�Լ�Ŀ�ͻȁ�Ŀ�Ć�ϵ�� ;(5) ��֧��ɳ�Ɔ�ϵ��;(6) ֧�ֻȁ�Ŀ�������AԪ�Ć�ϵ�ԣ��Ҹ������g��ϵ�y�l���Pϵ�飺( ���N���� + ( �ƿ��� + ( 䗛�쿂�� + ( �쿂�� + ( �L�ǻȿ��� + ( ţ�ȿ��� + �~��ȿ��� ) + ( �F�^�ȿ��� + �ȿ��� ))))))��
�����P�I�~ ��ֱ��Ŀ ;�����w����M ;���^����M ;�M��
����ֱ��Ŀ (Orthoptera) �`���ڹ�֫�����T (Arthropoda) ���x�V (Insecta)��Ŀǰȫ�����������ķN��� 27 941 �N (http://orthoptera.speciesfile.org/HomePage/ Orthoptera/HomePage.aspx)��ֱ��Ŀ���x��ռ�������������ϳ��O����������������Ϣ�أ������Bϵ�y�о��l�]����Ҫ�����ã��кܶ���N��������ʌW�����B�W�����ﻯ�W�������W�Լ��M������W���I���ģʽ���� [1-6]��Ŀǰ�����H���ձ��Jͬ��ֱ��Ŀ�֞�ȁ�Ŀ(Caelifera)��Ŀ(Ensifera)�������P��Ŀ�¸��AԪ�Ą�����Ȼ���ڷ��� [7]���ʴ_�b��ֱ��Ŀ���x�N������˽�ֱ��Ŀ���x����Դ�M����ϵ�y�l���Լ��NȺ�U������ֱ��Ŀ���x���ء������N�|�YԴ���þ�����Ҫ��ָ�����x��
�������x�����w����M��h���p朵Ĺ��r�]�� DNA ���ӣ��������������|��С���Y�����Ρ��߿�ؐ�����������������ء������M�����ʿ��Լ���N�Ȏ����l���ؽM��ĸϵ�z�����c��Ŀǰ�V�����������x��N�b��������ϵ�y�l���W����������W�Լ��NȺ�z���Y�����I����о� [8-11]����������S����ͨ���y��İlչ�������ˆT�@����Խ��Խ���ֱ��Ŀ���x�����w����M���У���Ļ���Mˮƽ���о�ֱ��Ŀ���x�M���ṩ�����õ����C��
�������ąR�����^ȥ 22 ���gֱ��Ŀ���x�����w����M�y��ijɹ��Լ����P�īI�����Y�����^�˻���M��С���A���M�ɡ��ܴa��ʹ�á������M�����ʡ��������ż�����Y����RNA �����ƅ^�Ļ���������������������ֱ��Ŀϵ�y�l���䣬��ϵ�y�о�ֱ��Ŀ���x�ķ��ϵ�y�l���e���˻��A�Y�ϡ�
����1 ֱ��Ŀ���x�����w����M�y��F��
����1995 �꣬Flook �� [12] �l���˵�һ��ֱ��Ŀ���x�����w�� (Locusta migratoria) �ľ����w����Mȫ���С��S���y���g�IJ���lչ������ 2017 �� 10 �� 13 �գ�GenBank �����칲��� 167 �Nֱ��Ŀ���xȫ�����w����M���С����Ё��Իȁ�Ŀ����N��࣬��Ӌ 99 �N���漰 9 ������ 22 ���� ;����Ŀ����N��Ӌ 68 �N���漰 7 ������ 12 ���ơ����б�����ҹ��ύ 53 �N (31.74%) ֱ��Ŀ���x�����w����M�����ؽ�ֱ��Ŀ���x�Ⱥ�gϵ�y�l���Pϵ����ʾֱ��Ŀ���x���M���^���ṩ���S���Ļ��A�����YԴ ( ���� 1)���mȻĿǰ�ѫ@��ȫ�L�����w����M����N�Ѹ��wֱ��Ŀ���x�����п��ƣ����c�����ֱ��Ŀ���x������ȣ��y�����N߀������������� Cooloolidae���]�� Mogoplistidae ��Ī��� Morabidae �Ȳ��ֿƼ��AԪ�Пo������N��
����2 ֱ��Ŀ���x�����w����M��������
����2.1�������w����M��С
�����c������������タ���w����M���ƣ�Ŀǰ�ќy�õ�����ֱ��Ŀ���xȫ�����w����M�����]�ϭh���p� DNA�������a 13 �������|���a����2 �� rRNA ����� 22 �� tRNA �����������w����M��С�Ğ� Hemicharilaus monomorphus[13]���L�Ȟ� 13 291 bp ;���ľ����w���G¶� (Phaneroptera gracilis)���L�Ȟ� 18 255 bp��ֱ��Ŀ���x�����w����M��С��׃������Ҫ�ܿ��ƅ^�����M�g��^�Ĕ������L��Ӱ푣��������^�� (Ruspolia dubia) [14] �Ŀ��ƅ^�L�ȃH�� 70 bp ;������M�L�����L���w���G¶���˾���һ���L�Ȟ� 1 548 bp �Ŀ��ƅ^���� nad2 �� trnW ֮�g߀����һ���L�Ȟ� 1 830 bp �Ļ����g��^��
����2.2�����������c����Y��
������������λ�ú��D䛷����׃�������x�����w�������ſɷ֞���λ ( ����λ�ø�׃���D䛷���׃ )����λ ( ����λ�ò�׃���D䛷����׃ ) �ͻ���ϴ�� ( ��������l����λ�� ( �� ) ��λ ) ���N��Ҫ����� [15]��Ŀǰ�ќy�����w����M��ֱ��Ŀ���x���� 8 �N�����w����M������� ���� 54 �Nֱ��Ŀ���x�ԈD 1-1 ��ʾ�ĵ��͵Ĺ�֫�����T�����w����M��ʽ���У����������Ŀ���x (50 �N ) �͘O�����Ļȁ�Ŀ���x (4 �N ) ;�� 90 �Nֱ��Ŀ���x�����w����M�� trnK �c trnD ����l����λ�����ԈD 1-2 ��ʾ�� trnD-trnK ��ʽ���� ( Ҳ�Q KD ���� )��ԓ�F��V�������ڳ����N���ƺ��쿂�Ƶ�׃ɫ���� (Erianthus versicolor) [13] ����Ļȁ�Ŀ���x�У���Ŀ���x���o KD ���ŬF��;�� 9 �Nֱ��Ŀ���x�����w����M�� trnN��trnS �c trnE ����l������ϴ�ƣ����ԈD 1-3 ��ʾ�� (-trnE)-(-trnS)-(-trnN)��(-trnE)- (trnS)-(-trnN) �� (-trnE)-( trnS)-(trnN) ���N��ʽ���ڣ�ԓ�F����������е������ ;�� 3 �Nֱ��Ŀ���x�����w����M�� trnI��trnQ �c trnM ����l������ϴ�ƣ����ԈD 1-4 ��ʾ�� trnM-trnI-(-trnQ) ��ʽ���ڣ�ԓ�F��H�����ڔM�~�x���� (Pseudophyllinae) �� ;�� 3 �Nֱ��Ŀ���x�����w����M�� AT �����^��trnI��trnQ��trnM �c nd2 ����l������ϴ�ƣ����ԈD 1-5 ��ʾ�� trnI-trnM-nd2-AT-(-trnQ) ��ʽ���ڣ�ԓ�F��H������¶�Ƶĸ��ϾG¶� (Holochlora fruhstorferi)���L���A�G¶� (Sinochlora longifissa) [16]���Ĵ��A�G¶� (Sinochlora szechwanensis) �� ;�Q�Ƶ� Cyphoderris monstrosa[6] �����w����M�� trnA �� trnR ����l����λ�����ԈD 1-6 ��ʾ�� trnR-trnA ��ʽ���� ;�˹�Ƶ����F�t� (Lipotactes tripyrga) �����w����M�� trnG��nd3��trnA��trnR��trnN �� trnS ����l������ϴ�ƣ����ԈD 1-7 ��ʾ�� trnRtrnS-trnA-trnN-trnG-nd3 �ķ�ʽ���� ;¶�Ƶ��۳���i¶� (Ruidocollaris obscura) [17] �����w����M�� AT �����^ͨ�^����ϴ�Ƴ��F�� trnY �� cox1 ֮�g�����ԈD 1-8 ��ʾ�� (-trnY)-AT-cox1 �ķ�ʽ���ڡ�
���������� 4 �N�ќyֱ��Ŀ���x�д�������ľ����w����M�Y�����քe���F�^�ȿƵĽ�� (Mekongiella kingdoni) [18] ȱ�� trnR ������ֺ�Ƶ� Comicus campestris[6] ȱ�� trnM �����ǻȿƵ� Locusta migratoria migratoria ����Ƶ� Troglophilus neglectus[19] �� cox1 �� cox2 ������� 2 �� trnL�����w���f��������Ĥ��Ŀ�ȏ��³�Ŀ���x [20]��ֱ��Ŀ���x�����w����M�����¼��������أ����˻ȁ�Ŀ�� KD ���ţ��H�� 23 �Nֱ��Ŀ���x�����w����M��������Y����l�������š�Ŀǰ�����������ŬF��Ľ����Ҫ�Џ��Ʒ��S�C�Gʧģ�͡������S�C�h��ģ�͡��ؽM�Լ��� tRNA �����e�`��ʼ����ď��� 4 �N��� [21]�����@Щģ���У������S�C�h��ģ���ѽ��@�ò����C����֧�֣���������Ȼ�]��һ�N������ģ�Ϳ��Խ�����е����ŬF�� [22]����ˣ����ڸ߶����ŵľ����w����M�����ö���ģ�ͽ�ጣ����磬���ڃH�l�� KD ���ŬF��Ļȁ�Ŀ�����Ï����S�C�h��ģ�ͽ�ጣ������ڰl���˻���ϴ�Ƶ�������͵ľ����w���ŬF�����Ҫ�Y�Ϗ��Ƅh��ģ�ͼ��ؽMģ�́���ͬ��ጡ�
����2.3���A���M���c�ܴa��ʹ����r
�����ќy��ֱ��Ŀ���xȫ�����w����M�������@�� AT ƫ���� ( ���� 2)��ƽ�� A+T ������ 73.31%������ A+T ������ߵĞ���Ƶ� Paramastax nigra[13] (78.00%)����͞郞���X� (Gampsocleis gratiosa) [23] (65.30%)��
������ֱ��Ŀ��Ŀ�������AԪ��ȫ�����w����M��ƽ�� AT �����������^�l�F ��2 ����Ŀ�У��ȁ�Ŀ�� A+T ���� (74.24%) ����Ŀ (71.95%) ;�ȁ�Ŀ�� 9 �������У��~��ȿ��Ƶ� A+T ������� (78.00%)�������N������� (68.80%) ;Ŀ�� 7 �������У������� A+T ������� (75.00%)����ԭ����� (69.45%)��һ����f��ͬһ�Ƽ��AԪ�е���N�g�A������׃���^С�������ڻȁ�Ŀ�У� A+T �����ڜy����N�����Ą��ǻȿ� (SE=0.12) �Ͱ]�ȿ� (SE=0.15) �гʬF�����@�ķ��B ;�����ǿ�����ˣ�������Ŀ�У��y����N�������˹�� (SE=0.48)��¶�� (SE=0.33) ����� Gryllidae (SE=0.63) �У���ͬ����N�g AT ������^�� ( �D 2)����ƵĬF��Ҳ���F�ڰ��Ŀ�� [22]��
���������ձ������� (Mecopoda niponensis) [24]�����ϾG¶� (Holochlora fruhstorferi) [11]���Ĵ��A�G¶� (Sinochlora szechwanensis)���ڽǾG¶� (Phaneroptera nigroantennata)��Tridactylus sp. �� Mirhipipteryx andensis[6] �⣬�����ќyֱ��Ŀ���x�����wȫ����M�� 4 �N�A���������� As>Ts>Cs>Gs��ͬ�r߀�ʬF���^���� A �A���� C �A��ƫ�� (AT-Skew = 0.11�� GC-Skew = -0.21)��ֱ��Ŀ���x�����w����M�ʬF���ĉA���M��ƫ�����c���M�����^����ͻ׃����Ȼ�x���S�C�z��Ư׃��ˮƽ�����D�ƺͻ���M�Y���ȶ�������������P�����¾����w����M�A������A>T��C>G���A���M��ƫ����A��C�ɷN�A��[25]������һ��������タ���w����M AT ��ƫ��� GC ؓƫ������c [26]��
������ֱ��Ŀ���x��Ŀ�������AԪ��ȫ�����w����M�A���M�ɵķ������^�l�F ( �D 3) ���ȁ�Ŀ��Ŀ�ĉA��ƫб�����Ϻ������タ���w����M���ձ�Ҏ�� ;�ȁ�Ŀ�ĉA���M��ƫб��� (AT skew = 0.15, GC skew = -0.18)��Ŀ��朵� AT �A���M�����ӽ� (AT skew = 0.05, GC skew = -0.27)���ȁ�Ŀ 9 �����ƵĉA��ƫб�������ձ�Ҏ�ɣ������N������朵�AT�A���M�����ӽ�(AT skew = 0.02)�⣬���� 8 ������ AT ƫб���������ң�AT ƫб������0.11~0.23;����䗛�쿂�ƵĉA���M��ƫб�(AT skew = 0.23, GC skew = -0.23)�����쿂�ƉA���M��ƫб���� (AT skew = 0.11, GC skew = -0.14)��Ŀ 7 �����ƵĉA��ƫб�������ձ�Ҏ�ɣ��� AT ƫб�������ڻȁ�Ŀ��AT ƫб������ 0.02~0.09 ;����ԭ�ƵĉA���M��ƫб��� (AT skew = 0.09, GC skew = -0.29)����� (AT skew = 0.08, GC skew = -0.28) ��֮���������� (AT skew = 0.02, GC skew = -0.24) ������
�����քe�yӋ 167 �N�ќyֱ��Ŀ���x�����w����M�����|���a������ܴa��ʹ����r��Ӌ�� 2 ����Ŀ������ͬ�x�ܴa�ӵ�ʹ���l�� (relative synonymous codon usage, RSCU)���yӋ�Y����D 4 ��ʾ �������ܴa���У�UUA(L) �ڻȁ�Ŀ��Ŀ�е� RSCU ֵ������ߣ��քe�� 2.91 �� 2.74 ;GCG(A) �ڻȁ�Ŀ��Ŀ�е� RSCU ֵ������ͣ��քe�� 0.11 �� 0.22 ;UGG(W) �� AUG(M) �ڻȁ�Ŀ��Ŀ�о��oƫ���� (RSCU = 1) ;�ܴa��ʹ��ƫ�����c�ܴa�ӵ���λ�c�� AT ƫ���ԳʬF��һ�����P�ԣ�����λ�c�� A �� U ���ܴa��ʹ���l���ձ��^�� ;ͬһ���ܴa�ӵ� RSCU ֵ��ֱ��Ŀ 2 ����Ŀ���x�g�o�@�����
����2.4��ֱ��Ŀ���x�����w�����|���a�����M������
����ֱ��Ŀ���x�� 13 �������w�����|���a����� A+T �������w���Ե������������w����M(����2)��
����������ƣ������|���a����ĵ�����λ�c A+T �����h�����ܴa�ӵ�һ����λ�c������y����N�����Ą��ǻȿ��ܴa�ӵĵ���λ�c A+T �������_ 76.38%�����ܴa�ӵ�һλ�c�͵ڶ�λ�c�H�� 73.85% �� 72.13%��
����ͨ�^���^��ֱ��Ŀ���x��ʼ�ܴa�Ӿ����������c ( ���D 1) ��atp6��cox2��cytB��nd3 �� nd4 ����ʼ�ܴa����˜ʵ����ܴa�� ATN (ATA��ATT��ATC�� ATG) ;cox3��atp8��nd1��nd2��nd4��nd5 �� nd6 ����ʼ�ܴa�ӳ��˘˜ʵ����ܴa�� ATN ���⣬߀���^�������ʼ�ܴa���� GTG��TTG��AGT��TTA�� CTG �� CCT ;���a cox1 �������ʼ�ܴa�������s�����˜ʵ����ܴa�� ATN ������ʼ�ܴa�ӣ��� cox1 ʹ���˷ǘ˜��ܴa�ӣ����� CCG��CAA�� ATT��TTA��ACC��ACG �ȣ����ⲻҎ�t�����ܴa�� ATGA[27]��GTGA[13]��ATAA[12] �ȶ��ǿ��ܵ� cox1 ����ʼ�ܴa�ӡ��@Щ�������ʼ�ܴa�ӿ��ԜpС�����g��^��ͬ�r�ֱ�����������l���دB��ͬ�r�����D䛳ɞ� mRNA ���^ RNA ���D�Q����������ʼ�ܴa�Ӳ���������ķ��g�^�� [22, 28]��
�����c��ʼ�ܴa�Ӳ�ͬ��ֱ��Ŀ���x�ڽKֹ�ܴa�ӷ�����F���^�ߵ�һ���� ( ���D 2) ��atp8 �ĽKֹ�ܴa�Ӟ���͵����ܴa�� TAA��TAG ;���� 12 �N�����|���a����ĽKֹ�ܴa�ӳ����������w�ܴa�� TAA �� TAG ֮�⣬߀�����������ĽKֹ�ܴa���� TA �� T���@�N�������ĽKֹ�ܴa���ں�������ľ����w����M�кܳ�Ҋ���о����Ɯy������D䛺�ͨ�^��������������γ������ĽKֹ�ܴa���M������D䛽Kֹ [29]��
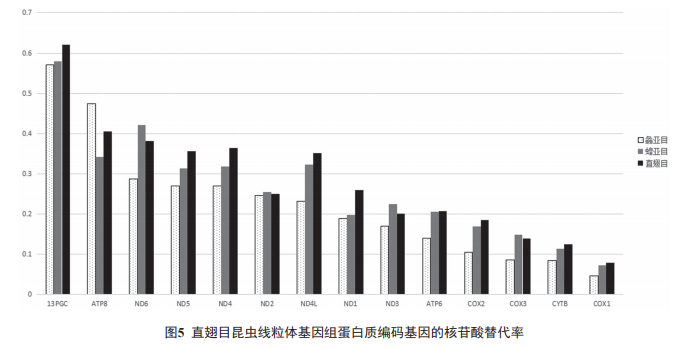
���������w����M��o�M���ױ��o�������ܵ����x���g�a����T׃�������ں˻���M�����^�ߵ��M������ [30]����ֱ��Ŀ 2 ����Ŀ�����w����M�ĵ����|���a�����M������ ( �÷�ͬ�x������cͬ�x����ʵı��� Ka/Ks ����ʾ�M������ ) �M�Ќ��ȣ������Y����D 5 ��ʾ ����ͬ�ĵ����|���a������M�����ʲ�ͬ������ atp8 �����M��������죬�� cox1 ��鱣�أ�13 �N�����|���a������ֱ��Ŀ�е��M���������� atp8 > nd6 > nd4 > nd5 > nd4L > nd1 > nd2 > atp6 > nd3 > cox2 > cox3 > cytB > cox1 ;��ͬ�ĵ����|���a�����ڲ�ͬ���Ⱥ��ͬ�Ӵ����@���IJ���� atp8 �������� 12 �������|���a�����ڻȁ�Ŀ�е��M�����ʾ�����Ŀ ;13 �������|���a����� Ka/Ks ��С�� 1�����F������ ( ؓ ) �x������l����ͬ�xͻ׃�������ӄݱ���̭�ˡ�
����2.5 tRNA��rRNA���c
�����ќy��ֱ��Ŀ���x�����w����M�У���� trnSAGN ȱ�� DHU �ۣ�ȱʧ DHU �ۺ�� trnSAGN �Կ��γɵ� L �������Y�����S�� CCA ���ܱ��c���ܴa���g�ľ��x [31] ;���� 21 �� tRNA �����ۯB�γɵ��͵����~�ݽY������ȻҲ�����⣬��ɽ�Ļ� (Sinopodisma houshana) �� trnP ֻ�� 58 bp����׃�h�� TΨC �۾������� [32]��tRNA ���γ����~�ݽY���A���䌦�r���l��һЩ�e�䣬��Ҫ�� G-U �e�䡣�@�N�F�������x�о��ձ���ڣ��ڌ�֩�� (Araneida) �����w tRNA ���о��аl�F���l���e��ĉA������ͨ�^�D䛺��֏������䌦���Ķ��Ɯy�����w tRNA ����ȱʧ DHU �ۻ� T �ی����������ܵ���ʹӰ푲��� [33]�����⣬ֱ��Ŀ���x tRNA �y���ķ��ܴa�ӷdz��̶�����δ�l�F��������ķ��ܴa�ӡ�
�������P֪�R���]�������w���Փ�İl��
����rRNA ������� rrnL ���� (lrRNA �� 16S rRNA)�� rrnS ���� (srRNA �� 12S rRNA)�����С��λ���������أ��҉A���M�ɱ��F�����@�� AT ƫ�á� rrnL ����Ķ����Y������ 6 ���Y���� (I��II��III�� IV��V �� VI)�����нY���� III �ڹ�֫���タ���w����M����ȱʧ�ģ��Y���� IV �� V ���س̶��^�� ; rrnS ����Ķ����Y�������Ă��Y���� (I��II��III �� IV)��ͨ���Y���� I �� II ׃���^�� III �� IV ���^���� [34]��ֱ��Ŀ���x�����w����M�� rrnL ����λ�� trnLCUN �� trnV ֮�g��ƽ���L�Ȟ� 1 317 bp���������L�Ğ���dz�� (Calliptamus abbreviatus)���L�Ȟ� 1 555 bp ;��̵Ğ�W���N�� (Gryllotalpa pluvialis)���L�Ȟ� 1 236 bp[19]��ֱ��Ŀ���x�����w����M rrnS ������L���hС�� rrnL ����ƽ���L�ȃH�� 800 bp���������L�Ğ����Ƶ� Trigonidium sjostedti���L�Ȟ� 1 004 bp[35]������̵Ğ鰵ɫ��� (Phlaeoba tenebrosa)���L�ȃH�� 212 bp��
����2.6�����ƅ^���c
����ֱ��Ŀ���x���ƅ^λ���������أ�ͨ��λ�� rrnS �� trnI ֮�g����Ҳ�����⣬���縵�ϾG¶�L���A�G¶���Ĵ��A�G¶�Ŀ��ƅ^λ�� nd2 �� trnQ ֮�g������i¶�Ŀ��ƅ^λ�� trnY �� cox1 ֮�g ;�L�Ȳ�ܴ�������̵Ğ����^�� [14]���H�� 70 bp�������L�Ğ��L���A�G¶� [16]�����ƅ^�L�Ȟ� 3 122 bp����ͬ��N�g���ƅ^���L��׃����Ҫ�c�߶��؏͵����еĔ����ʹ�С���P���@Щ�؏����п��Mһ���γɺ��εİl�A�Y���������s�Ķ����Y������K���Ƶ��M�̡����о��������M�ܲ�ͬ��N�����w N 朵ď������cλ�ò�ͬ����һЩȫ׃�B���x�ľ����w�д��ڵ� T-strech (ploy-T > 10 bp) �����c������ʼ�R�e���P�������ְ�׃�B���x�ľ����w����M������ T-strech �Y�� [36]����ֱ��Ŀ�У��ֻȁ�Ŀ�Լ�Ŀ����ƺ��N���������x�ľ����w���ƅ^���mȻ�]�� T-strech �Y�������о������䌦��λ�ðl�F ploy-T �ĽY�����A�� C ��࣬���γ����������o�h�Y������������w��(Locusta migratoria) [12]���؎X�Ļ� (Sinopodisma tsinlingensis) [37]��ɮñ��� (Phlaeoba infumata) [37]��С�~�� (Traulia minuta) [37] �� ;��Ŀ���˹���Ǝ����y�A�y���@�ӵ��o�h�Y�� [34]�����о������˹���Ʋ�����N�ľ����w���ƅ^ N ��ϰl�F�� T-strech �Y��������Ħ�T�˹ (Anabrus simplex) [38]�������i� (Deracantha onos) [39]�������X� (Gampsocleis gratiosa) [38] �ͺڽǾG¶� (Phaneroptera nigroantennata) [37]��������֮�����x�����w���ƅ^�������L�ȺͽY��׃���ܴ�——Փ�����ߣ��w����1,2����ѩ��1 ���S��ԭ1 *
SCISSCIAHCI